| [1] |
Barratt C L R, Björndahl L, De Jonge C J, et al. The diagnosis of male infertility: an analysis of the evidence to support the development of global WHO guidance—challenges and future research opportunities. Human Reproduction Update, 2017, 23 (6): 660–680. doi: 10.1093/humupd/dmx021
|
| [2] |
Ma Y J, Wu B B, Chen Y H, et al. CCDC146 is required for sperm flagellum biogenesis and male fertility in mice. Cellular and Molecular Life Sciences, 2024, 81: 1. doi: 10.1007/s00018-023-05025-x
|
| [3] |
Long S H, Fu L L, Ma J, et al. Novel biallelic variants in DNAH1 cause multiple morphological abnormalities of sperm flagella with favorable outcomes of fertility after ICSI in Han Chinese males. Andrology, 2024, 12 (2): 349–364. doi: 10.1111/andr.13476
|
| [4] |
Zhang B B, Khan I, Liu C Y, et al. Novel loss-of-function variants in DNAH17 cause multiple morphological abnormalities of the sperm flagella in humans and mice. Clinical Genetics, 2021, 99 (1): 176–186. doi: 10.1111/cge.13866
|
| [5] |
Meng X M, Xu C, Li J W, et al. Multi-scale structures of the mammalian radial spoke and divergence of axonemal complexes in ependymal cilia. Nature Communications, 2024, 15: 362. doi: 10.1038/s41467-023-44577-1
|
| [6] |
Zhang G H, Li D Y, Tu C F, et al. Loss-of-function missense variant of AKAP4 induced male infertility through reduced interaction with QRICH2 during sperm flagella development. Human Molecular Genetics, 2022, 31 (2): 219–231. doi: 10.1093/hmg/ddab234
|
| [7] |
Visser L, Westerveld G H, Xie F, et al. A comprehensive gene mutation screen in men with asthenozoospermia. Fertility and Sterility, 2011, 95 (3): 1020–1024.e9. doi: 10.1016/j.fertnstert.2010.11.067
|
| [8] |
Wu B B, Yu X C, Liu C, et al. Essential role of CFAP53 in sperm flagellum biogenesis. Frontiers in Cell and Developmental Biology, 2021, 9: 676910. doi: 10.3389/fcell.2021.676910
|
| [9] |
Xu K B, Yang L L, Zhang L, et al. Lack of AKAP3 disrupts integrity of the subcellular structure and proteome of mouse sperm and causes male sterility. Development, 2020, 147 (2): dev181057. doi: 10.1242/dev.181057
|
| [10] |
Xu C, Tang D D, Shao Z M, et al. Homozygous SPAG6 variants can induce nonsyndromic asthenoteratozoospermia with severe MMAF. Reproductive Biology and Endocrinology, 2022, 20: 41. doi: 10.1186/s12958-022-00916-3
|
| [11] |
Xu Y J, Yang B Y, Lei C, et al. Novel compound heterozygous variants in CCDC40 associated with primary ciliary dyskinesia and multiple morphological abnormalities of the sperm flagella. Pharmacogenomics and Personalized Medicine, 2022, 15: 341–350. doi: 10.2147/PGPM.S359821
|
| [12] |
Yin Y Y, Mu W Y, Yu X C, et al. LRRC46 accumulates at the midpiece of sperm flagella and is essential for spermiogenesis and male fertility in mouse. International Journal of Molecular Sciences, 2022, 23 (15): 8525. doi: 10.3390/ijms23158525
|
| [13] |
Zhang J T, He X J, Wu H, et al. Loss of DRC1 function leads to multiple morphological abnormalities of the sperm flagella and male infertility in human and mouse. Human Molecular Genetics, 2021, 30 (21): 1996–2011. doi: 10.1093/hmg/ddab171
|
| [14] |
Zhang R D, Wu B B, Liu C, et al. CCDC38 is required for sperm flagellum biogenesis and male fertility in mice. Development, 2022, 149 (11): dev200516. doi: 10.1242/dev.200516
|
| [15] |
Ito C, Akutsu H, Yao R, et al. Odf2 haploinsufficiency causes a new type of decapitated and decaudated spermatozoa, Odf2-DDS, in mice. Scientific Reports, 2019, 9: 14249. doi: 10.1038/s41598-019-50516-2
|
| [16] |
Khelifa M B, Coutton C, Zouari R, et al. Mutations in DNAH1, which encodes an inner arm heavy chain dynein, lead to male infertility from multiple morphological abnormalities of the sperm flagella. The American Journal of Human Genetics, 2014, 94 (1): 95–104. doi: 10.1016/j.ajhg.2013.11.017
|
| [17] |
Chang T L, Tang H Y, Zhou X, et al. A novel homozygous nonsense variant of AK7 is associated with multiple morphological abnormalities of the sperm flagella. Reproductive Biomedicine Online, 2024, 48 (5): 103765. doi: 10.1016/j.rbmo.2023.103765
|
| [18] |
Gu L J, Liu X M, Yang J, et al. A new hemizygous missense mutation, c.454T>C (p.S152P), in AKAP4 gene is associated with asthenozoospermia. Molecular Reproduction and Development, 2021, 88 (9): 587–597. doi: 10.1002/mrd.23529
|
| [19] |
Liu C Y, Shen Y, Tang S Y, et al. Homozygous variants in AKAP3 induce asthenoteratozoospermia and male infertility. Journal of Medical Genetics, 2023, 60 (2): 137–143. doi: 10.1136/jmedgenet-2021-108271
|
| [20] |
Wang M, Yang Q Y, Zhou J P, et al. Novel compound heterozygous mutations in DNAH1 cause primary infertility in Han Chinese males with multiple morphological abnormalities of the sperm flagella. Asian Journal of Andrology, 2023, 25 (4): 512–519. doi: 10.4103/aja202292
|
| [21] |
Zhu Z J, Wang Y Z, Wang X B, et al. Novel mutation in ODF2 causes multiple morphological abnormalities of the sperm flagella in an infertile male. Asian Journal of Andrology, 2022, 24 (5): 463–472. doi: 10.4103/aja202183
|
| [22] |
Shen Y, Zhang F, Li F P, et al. Loss-of-function mutations in QRICH2 cause male infertility with multiple morphological abnormalities of the sperm flagella. Nature Communications, 2019, 10: 433. doi: 10.1038/s41467-018-08182-x
|
| [23] |
Kherraf Z E, Cazin C, Coutton C, et al. Whole exome sequencing of men with multiple morphological abnormalities of the sperm flagella reveals novel homozygous QRICH2 mutations. Clinical Genetics, 2019, 96 (5): 394–401. doi: 10.1111/cge.13604
|
| [24] |
Hiltpold M, Janett F, Mapel X M, et al. A 1-bp deletion in bovine QRICH2 causes low sperm count and immotile sperm with multiple morphological abnormalities. Genetics Selection Evolution, 2022, 54: 18. doi: 10.1186/s12711-022-00710-0
|
| [25] |
Ullah M A, Husseni A M, Mahmood S U. Consanguineous marriages and their detrimental outcomes in Pakistan: an urgent need for appropriate measures. International Journal of Community Medicine and Public Health, 2017, 5 (1): 1–3. doi: 10.18203/2394-6040.ijcmph20175757
|
| [26] |
Björndahl L, Brown J K. The sixth edition of the WHO Laboratory Manual for the Examination and Processing of Human Semen: ensuring quality and standardization in basic examination of human ejaculates. Fertility and Sterility, 2022, 117 (2): 246–251. doi: 10.1016/j.fertnstert.2021.12.012
|
| [27] |
Li H, Durbin R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics, 2009, 25 (14): 1754–1760. doi: 10.1093/bioinformatics/btp324
|
| [28] |
Wang K, Li M Y, Hakonarson H. ANNOVAR: functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Research, 2010, 38 (16): e164. doi: 10.1093/nar/gkq603
|
| [29] |
McKenna A, Hanna M, Banks E, et al. The Genome Analysis Toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Research, 2010, 20 (9): 1297–1303. doi: 10.1101/gr.107524.110
|
| [30] |
Quinodoz M, Peter V G, Bedoni N, et al. AutoMap is a high performance homozygosity mapping tool using next-generation sequencing data. Nature Communications, 2021, 12: 518. doi: 10.1038/s41467-020-20584-4
|
| [31] |
Pedersen B S, Quinlan A R. Who’s who? Detecting and resolving sample anomalies in human DNA sequencing studies with peddy. The American Journal of Human Genetics, 2017, 100 (3): 406–413. doi: 10.1016/j.ajhg.2017.01.017
|
| [32] |
Zhang B B, Ma H, Khan T, et al. A DNAH17 missense variant causes flagella destabilization and asthenozoospermia. Journal of Experimental Medicine, 2020, 217 (2): e20182365. doi: 10.1084/jem.20182365
|
| [33] |
Ma A, Zeb A, Ali I, et al. Biallelic variants in CFAP61 cause multiple morphological abnormalities of the flagella and male infertility. Frontiers in Cell and Developmental Biology, 2022, 9: 803818. doi: 10.3389/fcell.2021.803818
|
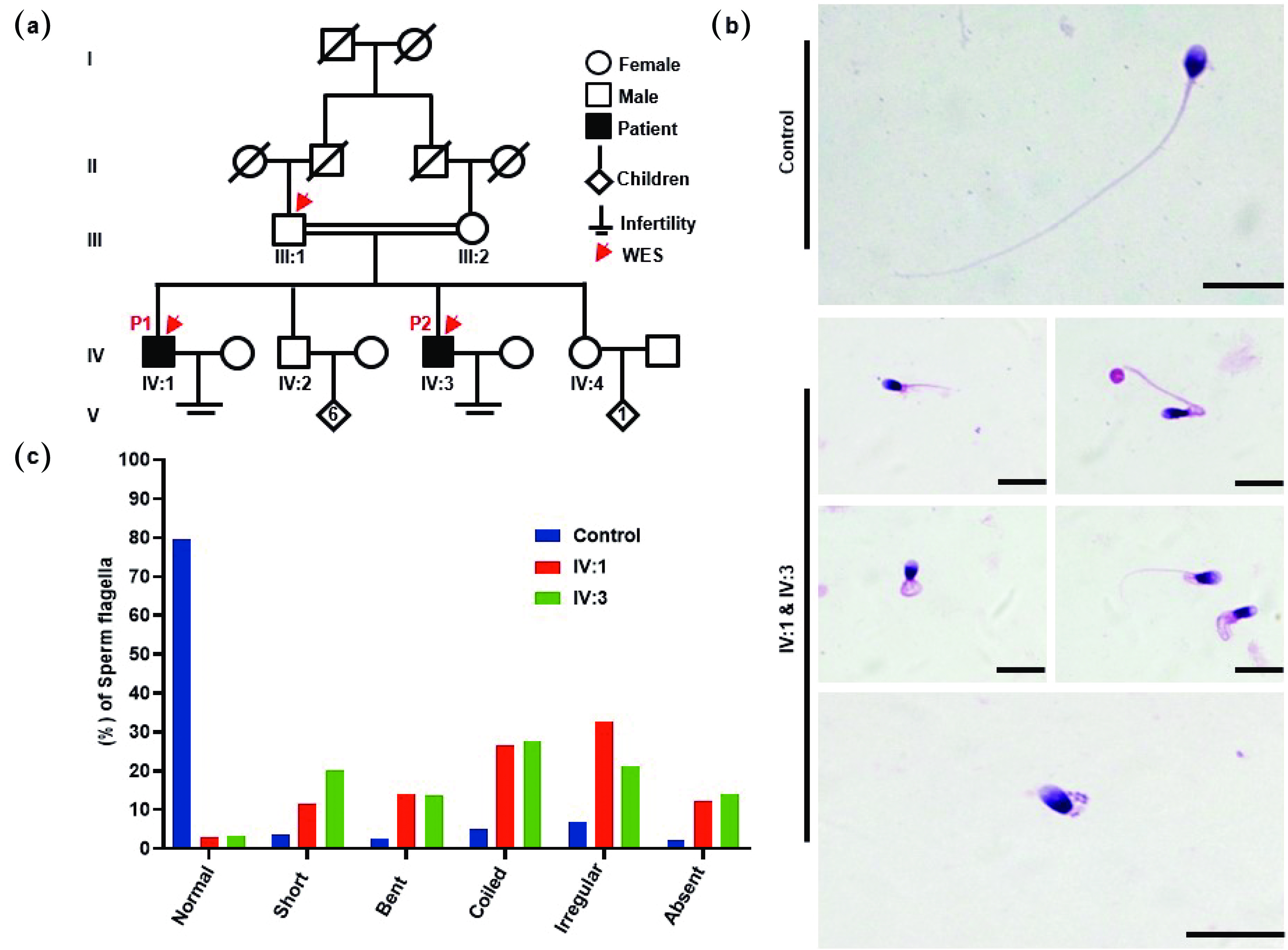
Figure 1. MMAF patients from a Pakistani consanguineous family. (a) Pedigree chart of two infertile male patients, IV:1 and IV:3, from a consanguineous marriage. Squares denote males, while circles represent females. The solid squares denote the patients, and the hollow squares represent the unaffected individuals. The double horizontal lines indicate a consanguineous marriage. The Arabic numerals indicate the number of children born to a couple, whereas the Roman numerals indicate the generation number. The red arrows on the pedigree chart represent individuals analyzed via WES. (b) H&E staining showing the morphological defects of flagella in the spermatozoa of MMAF patients, including (i) short, (ii) bent, (iii) coiled, (iv) irregular caliber, and (v) absent. (c) Statistics of flagellar abnormalities in both the fertile controls and the patients (IV:1 & IV:3). Scale bars = 10 µm. WES: whole-exome sequencing.
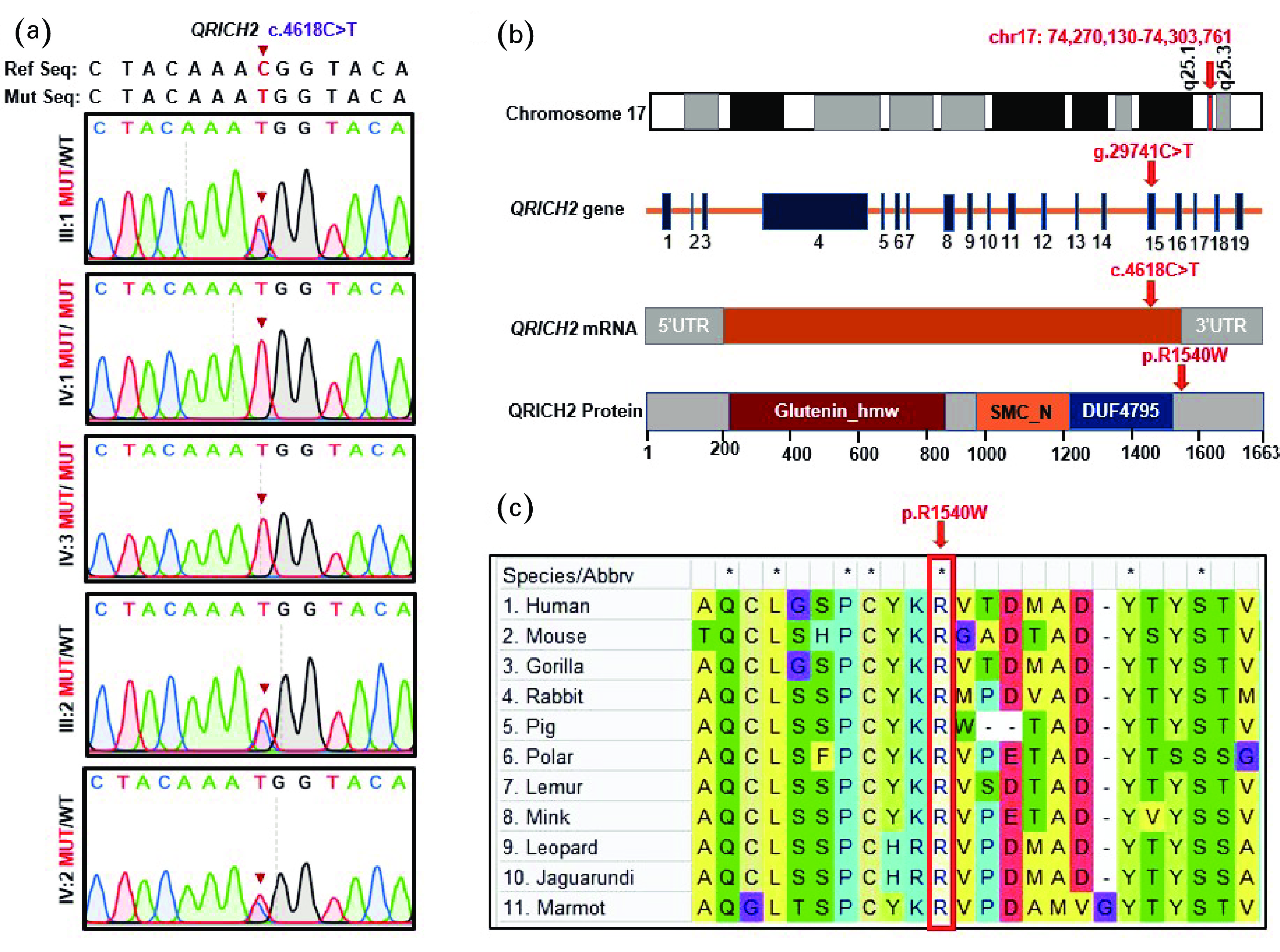
Figure
2.
Identification of a novel homozygous QRICH2 missense variant. (a) Sanger sequencing verification of the QRICH2 mutation, c.4618C>T, across available family members. The parents (III:1 & III:2) and the fertile brother (IV:2) were heterozygous, whereas the patients (IV:1 & IV:3) were homozygous for the identified mutation. Red arrows indicate the identified mutation. (b) QRICH2 structure and position of the identified mutation at the genomic, transcriptional, and protein levels. QRICH2 is located at chromosome 17 and consists of 19 exons encoding a protein of
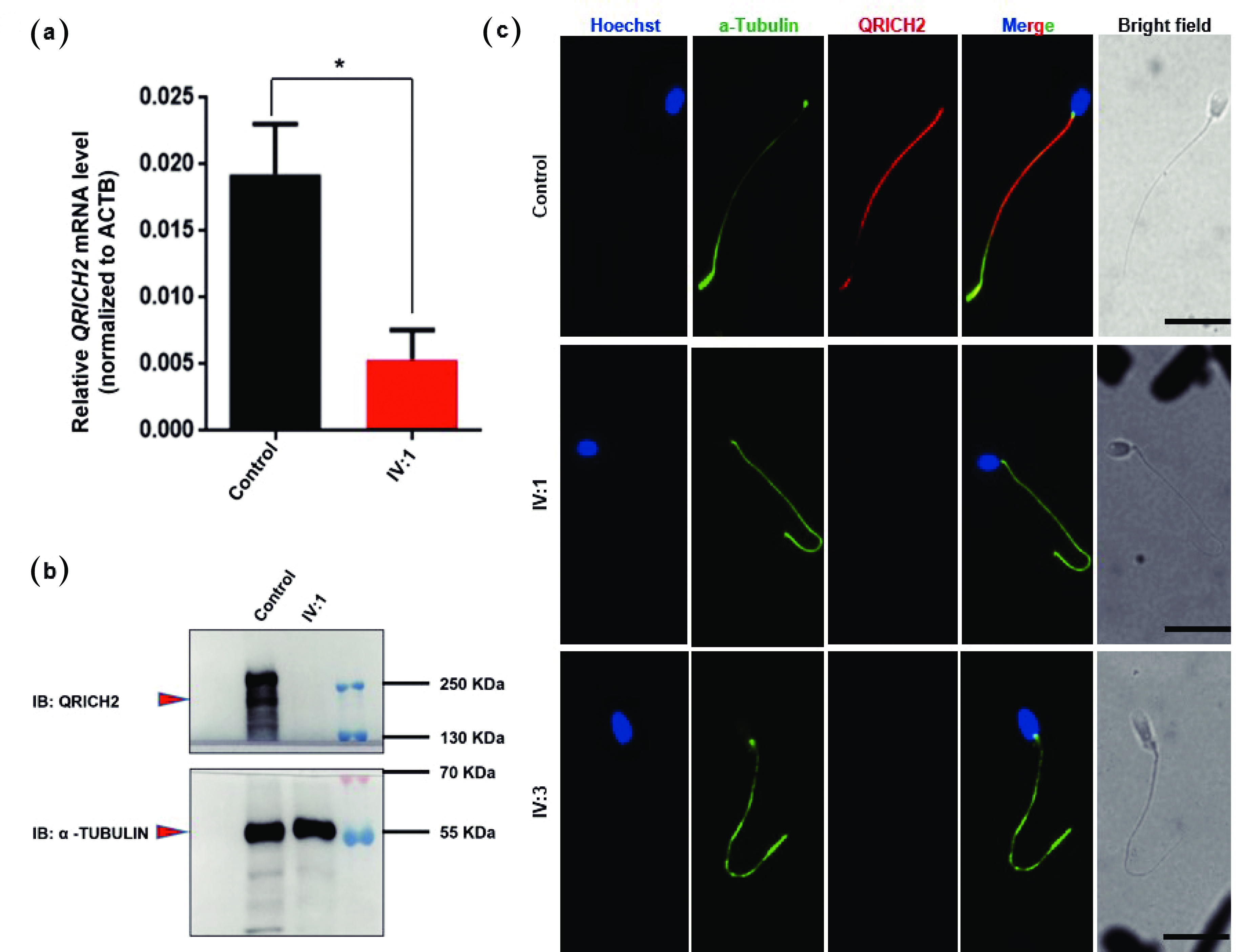
Figure 3. QRICH2 expression was absent in the patient’s spermatozoa. (a) The graph represents the QRICH2 mRNA expression level of patient IV:1 with a low level of QRICH2 compared with that of the fertile control. n = 3, Student’s t test; *P < 0.05. (b) Representative western blot images showing the presence of the QRICH2 band in the sperm lysate of the normal fertile control and the complete absence of the band in patient IV:1. A loading control (e.g., α-tubulin) was used to ensure equal protein loading. (c) Representative images of spermatozoa from normal fertile controls and patients (IV:1 & IV:3) costained with an anti-QRICH2 antibody (red), an anti-α-tubulin antibody (green), and Hoechst (blue, nuclear marker). The QRICH2 signals were absent in the spermatozoa of the patients (IV:1 & IV:3), whereas normal signals of QRICH2 were observed in the anterior sperm flagella of the normal fertile controls. Scale bars = 10 µm. QRICH2: Glutamine-rich protein 2. Statistical analysis revealed that the difference in QRICH2 expression between patients (IV:1) and normal fertile controls was significant (*P<0.05).
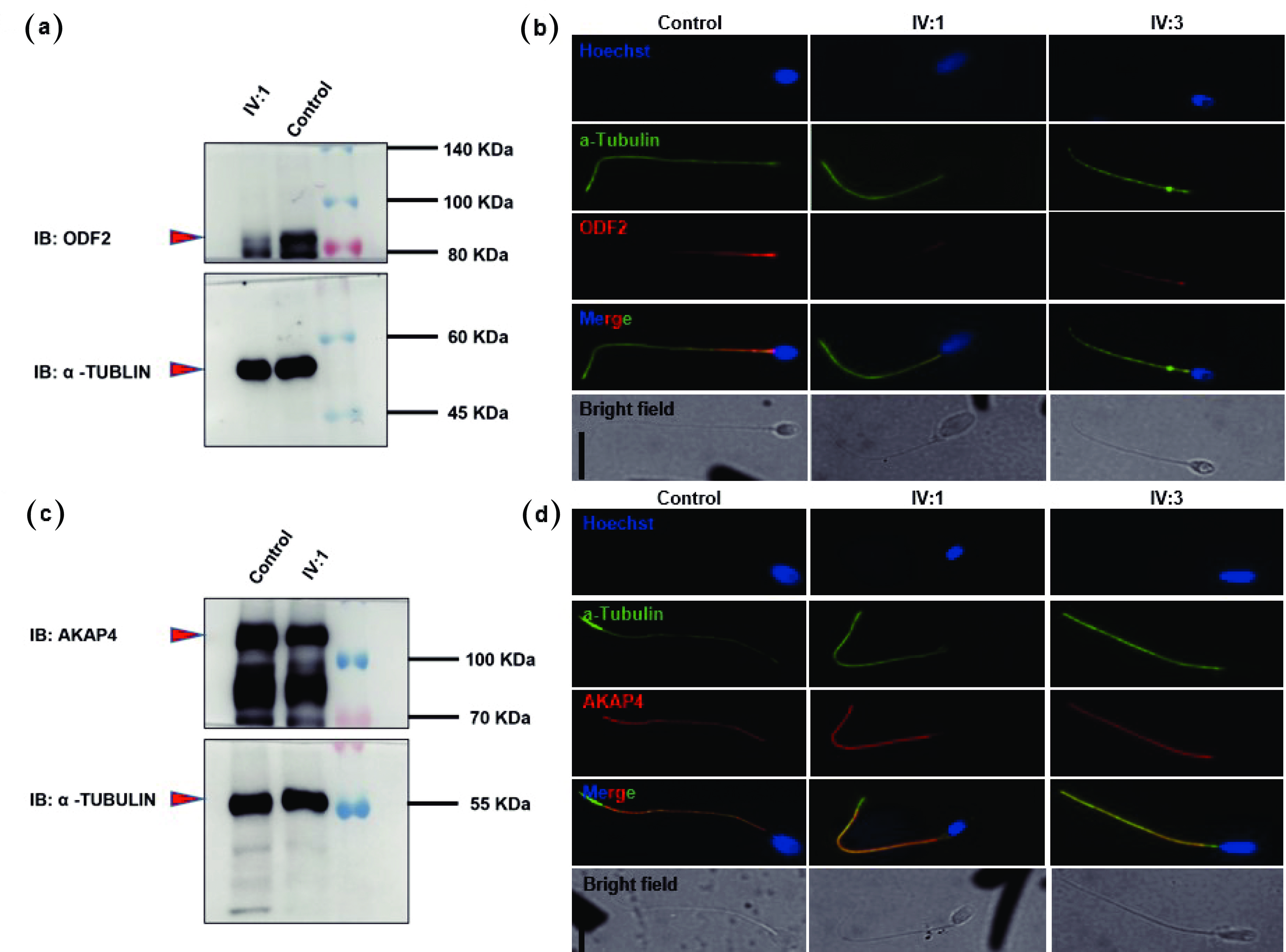
Figure 4. Patients with the QRICH2 mutation presented reduced ODF2 and normal AKAP4 expression in their spermatozoa. Western blot analysis revealed the presence of a weak band of ODF2 (a) and an intact band of AKAP4 (c) in the sperm lysate of patient IV:1. Spermatozoa from normal fertile controls and patients (IV:1 & IV:3) were costained with anti-ODF2 and anti-AKAP4 antibodies. Images (b) and (d) show the corresponding fluorescence microscopy images of spermatozoa stained with anti-ODF2 and anti-AKAP4 antibodies, an anti-α-tubulin antibody (green), and Hoechst (a nuclear marker). The signals of ODF2 were weak (b), whereas normal signals of AKAP4 were detected in the spermatozoa of the patients (IV:1 & IV:3) (d). Scale bars = 10 µm. QRICH2: Glutamine-rich protein 2. ODF2: Outer dense fiber of sperm tails 2. AKAP4: A-kinase anchoring protein 4.
| [1] |
Barratt C L R, Björndahl L, De Jonge C J, et al. The diagnosis of male infertility: an analysis of the evidence to support the development of global WHO guidance—challenges and future research opportunities. Human Reproduction Update, 2017, 23 (6): 660–680. doi: 10.1093/humupd/dmx021
|
| [2] |
Ma Y J, Wu B B, Chen Y H, et al. CCDC146 is required for sperm flagellum biogenesis and male fertility in mice. Cellular and Molecular Life Sciences, 2024, 81: 1. doi: 10.1007/s00018-023-05025-x
|
| [3] |
Long S H, Fu L L, Ma J, et al. Novel biallelic variants in DNAH1 cause multiple morphological abnormalities of sperm flagella with favorable outcomes of fertility after ICSI in Han Chinese males. Andrology, 2024, 12 (2): 349–364. doi: 10.1111/andr.13476
|
| [4] |
Zhang B B, Khan I, Liu C Y, et al. Novel loss-of-function variants in DNAH17 cause multiple morphological abnormalities of the sperm flagella in humans and mice. Clinical Genetics, 2021, 99 (1): 176–186. doi: 10.1111/cge.13866
|
| [5] |
Meng X M, Xu C, Li J W, et al. Multi-scale structures of the mammalian radial spoke and divergence of axonemal complexes in ependymal cilia. Nature Communications, 2024, 15: 362. doi: 10.1038/s41467-023-44577-1
|
| [6] |
Zhang G H, Li D Y, Tu C F, et al. Loss-of-function missense variant of AKAP4 induced male infertility through reduced interaction with QRICH2 during sperm flagella development. Human Molecular Genetics, 2022, 31 (2): 219–231. doi: 10.1093/hmg/ddab234
|
| [7] |
Visser L, Westerveld G H, Xie F, et al. A comprehensive gene mutation screen in men with asthenozoospermia. Fertility and Sterility, 2011, 95 (3): 1020–1024.e9. doi: 10.1016/j.fertnstert.2010.11.067
|
| [8] |
Wu B B, Yu X C, Liu C, et al. Essential role of CFAP53 in sperm flagellum biogenesis. Frontiers in Cell and Developmental Biology, 2021, 9: 676910. doi: 10.3389/fcell.2021.676910
|
| [9] |
Xu K B, Yang L L, Zhang L, et al. Lack of AKAP3 disrupts integrity of the subcellular structure and proteome of mouse sperm and causes male sterility. Development, 2020, 147 (2): dev181057. doi: 10.1242/dev.181057
|
| [10] |
Xu C, Tang D D, Shao Z M, et al. Homozygous SPAG6 variants can induce nonsyndromic asthenoteratozoospermia with severe MMAF. Reproductive Biology and Endocrinology, 2022, 20: 41. doi: 10.1186/s12958-022-00916-3
|
| [11] |
Xu Y J, Yang B Y, Lei C, et al. Novel compound heterozygous variants in CCDC40 associated with primary ciliary dyskinesia and multiple morphological abnormalities of the sperm flagella. Pharmacogenomics and Personalized Medicine, 2022, 15: 341–350. doi: 10.2147/PGPM.S359821
|
| [12] |
Yin Y Y, Mu W Y, Yu X C, et al. LRRC46 accumulates at the midpiece of sperm flagella and is essential for spermiogenesis and male fertility in mouse. International Journal of Molecular Sciences, 2022, 23 (15): 8525. doi: 10.3390/ijms23158525
|
| [13] |
Zhang J T, He X J, Wu H, et al. Loss of DRC1 function leads to multiple morphological abnormalities of the sperm flagella and male infertility in human and mouse. Human Molecular Genetics, 2021, 30 (21): 1996–2011. doi: 10.1093/hmg/ddab171
|
| [14] |
Zhang R D, Wu B B, Liu C, et al. CCDC38 is required for sperm flagellum biogenesis and male fertility in mice. Development, 2022, 149 (11): dev200516. doi: 10.1242/dev.200516
|
| [15] |
Ito C, Akutsu H, Yao R, et al. Odf2 haploinsufficiency causes a new type of decapitated and decaudated spermatozoa, Odf2-DDS, in mice. Scientific Reports, 2019, 9: 14249. doi: 10.1038/s41598-019-50516-2
|
| [16] |
Khelifa M B, Coutton C, Zouari R, et al. Mutations in DNAH1, which encodes an inner arm heavy chain dynein, lead to male infertility from multiple morphological abnormalities of the sperm flagella. The American Journal of Human Genetics, 2014, 94 (1): 95–104. doi: 10.1016/j.ajhg.2013.11.017
|
| [17] |
Chang T L, Tang H Y, Zhou X, et al. A novel homozygous nonsense variant of AK7 is associated with multiple morphological abnormalities of the sperm flagella. Reproductive Biomedicine Online, 2024, 48 (5): 103765. doi: 10.1016/j.rbmo.2023.103765
|
| [18] |
Gu L J, Liu X M, Yang J, et al. A new hemizygous missense mutation, c.454T>C (p.S152P), in AKAP4 gene is associated with asthenozoospermia. Molecular Reproduction and Development, 2021, 88 (9): 587–597. doi: 10.1002/mrd.23529
|
| [19] |
Liu C Y, Shen Y, Tang S Y, et al. Homozygous variants in AKAP3 induce asthenoteratozoospermia and male infertility. Journal of Medical Genetics, 2023, 60 (2): 137–143. doi: 10.1136/jmedgenet-2021-108271
|
| [20] |
Wang M, Yang Q Y, Zhou J P, et al. Novel compound heterozygous mutations in DNAH1 cause primary infertility in Han Chinese males with multiple morphological abnormalities of the sperm flagella. Asian Journal of Andrology, 2023, 25 (4): 512–519. doi: 10.4103/aja202292
|
| [21] |
Zhu Z J, Wang Y Z, Wang X B, et al. Novel mutation in ODF2 causes multiple morphological abnormalities of the sperm flagella in an infertile male. Asian Journal of Andrology, 2022, 24 (5): 463–472. doi: 10.4103/aja202183
|
| [22] |
Shen Y, Zhang F, Li F P, et al. Loss-of-function mutations in QRICH2 cause male infertility with multiple morphological abnormalities of the sperm flagella. Nature Communications, 2019, 10: 433. doi: 10.1038/s41467-018-08182-x
|
| [23] |
Kherraf Z E, Cazin C, Coutton C, et al. Whole exome sequencing of men with multiple morphological abnormalities of the sperm flagella reveals novel homozygous QRICH2 mutations. Clinical Genetics, 2019, 96 (5): 394–401. doi: 10.1111/cge.13604
|
| [24] |
Hiltpold M, Janett F, Mapel X M, et al. A 1-bp deletion in bovine QRICH2 causes low sperm count and immotile sperm with multiple morphological abnormalities. Genetics Selection Evolution, 2022, 54: 18. doi: 10.1186/s12711-022-00710-0
|
| [25] |
Ullah M A, Husseni A M, Mahmood S U. Consanguineous marriages and their detrimental outcomes in Pakistan: an urgent need for appropriate measures. International Journal of Community Medicine and Public Health, 2017, 5 (1): 1–3. doi: 10.18203/2394-6040.ijcmph20175757
|
| [26] |
Björndahl L, Brown J K. The sixth edition of the WHO Laboratory Manual for the Examination and Processing of Human Semen: ensuring quality and standardization in basic examination of human ejaculates. Fertility and Sterility, 2022, 117 (2): 246–251. doi: 10.1016/j.fertnstert.2021.12.012
|
| [27] |
Li H, Durbin R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics, 2009, 25 (14): 1754–1760. doi: 10.1093/bioinformatics/btp324
|
| [28] |
Wang K, Li M Y, Hakonarson H. ANNOVAR: functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Research, 2010, 38 (16): e164. doi: 10.1093/nar/gkq603
|
| [29] |
McKenna A, Hanna M, Banks E, et al. The Genome Analysis Toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Research, 2010, 20 (9): 1297–1303. doi: 10.1101/gr.107524.110
|
| [30] |
Quinodoz M, Peter V G, Bedoni N, et al. AutoMap is a high performance homozygosity mapping tool using next-generation sequencing data. Nature Communications, 2021, 12: 518. doi: 10.1038/s41467-020-20584-4
|
| [31] |
Pedersen B S, Quinlan A R. Who’s who? Detecting and resolving sample anomalies in human DNA sequencing studies with peddy. The American Journal of Human Genetics, 2017, 100 (3): 406–413. doi: 10.1016/j.ajhg.2017.01.017
|
| [32] |
Zhang B B, Ma H, Khan T, et al. A DNAH17 missense variant causes flagella destabilization and asthenozoospermia. Journal of Experimental Medicine, 2020, 217 (2): e20182365. doi: 10.1084/jem.20182365
|
| [33] |
Ma A, Zeb A, Ali I, et al. Biallelic variants in CFAP61 cause multiple morphological abnormalities of the flagella and male infertility. Frontiers in Cell and Developmental Biology, 2022, 9: 803818. doi: 10.3389/fcell.2021.803818
|

ISSN 0253-2778
CN 34-1054/N
Copyright © Editorial Office of JUSTC, All Rights Reserved. 皖ICP备05002528号
Supported by:
Beijing Renhe Information Technology Co. Ltd





 DownLoad:
DownLoad: