| [1] |
Dixit D, Okuniewska M, Schwab S R. Secrets and lyase: Control of sphingosine 1-phosphate distribution. Immunol. Rev., 2019, 289 (1): 173–185. doi: 10.1111/imr.12760
|
| [2] |
Książek M, Chacińska M, Chabowski A, et al. Sources, metabolism, and regulation of circulating sphingosine-1-phosphate. J. Lipid Res., 2015, 56 (7): 1271–1281. doi: 10.1194/jlr.R059543
|
| [3] |
Spiegel S, Milstien S. Sphingosine-1-phosphate: an enigmatic signalling lipid. Nat. Rev. Mol. Cell Biol., 2003, 4 (5): 397–407. doi: 10.1038/nrm1103
|
| [4] |
Nagahashi M, Takabe K, Terracina K P, et al. Sphingosine-1-phosphate transporters as targets for cancer therapy. Biomed Res. Int., 2014, 2014: 651727. doi: 10.1155/2014/651727
|
| [5] |
Chun J, Hla T, Lynch K R, et al. International Union of Basic and Clinical Pharmacology. LXXVIII. Lysophospholipid receptor nomenclature. Pharmacol. Rev., 2010, 62 (4): 579–587. doi: 10.1124/pr.110.003111
|
| [6] |
Hait N C, Oskeritzian C A, Paugh S W, et al. Sphingosine kinases, sphingosine 1-phosphate, apoptosis and diseases. Biochim. Biophys. Acta, 2006, 1758 (12): 2016–2026. doi: 10.1016/j.bbamem.2006.08.007
|
| [7] |
Ng M L, Yarla N S, Menschikowski M, et al. Regulatory role of sphingosine kinase and sphingosine-1-phosphate receptor signaling in progenitor/stem cells. World J. Stem Cells, 2018, 10 (9): 119–133. doi: 10.4252/wjsc.v10.i9.119
|
| [8] |
Sukocheva O A. Expansion of sphingosine kinase and sphingosine-1-phosphate receptor function in normal and cancer cells: From membrane restructuring to mediation of estrogen signaling and stem cell programming. Int. J. Mol. Sci., 2018, 19 (2): 420. doi: 10.3390/ijms19020420
|
| [9] |
Birchwood C J, Saba J D, Dickson R C, et al. Calcium influx and signaling in yeast stimulated by intracellular sphingosine 1-phosphate accumulation. J. Biol. Chem., 2001, 276 (15): 11712–11718. doi: 10.1074/jbc.M010221200
|
| [10] |
Pulli I, Asghar M Y, Kemppainen K, et al. Sphingolipid-mediated calcium signaling and its pathological effects. Biochim. Biophys. Acta Mol. Cell Res., 2018, 1865 (11 Pt B): 1668–1677. doi: 10.1016/j.bbamcr.2018.04.012
|
| [11] |
Putney J W, Tomita T. Phospholipase C signaling and calcium influx. Adv. Biol. Regul., 2012, 52 (1): 152–164. doi: 10.1016/j.advenzreg.2011.09.005
|
| [12] |
Meyer zu Heringdorf D, Liliom K, Schaefer M, et al. Photolysis of intracellular caged sphingosine-1-phosphate causes Ca2+ mobilization independently of G-protein-coupled receptors. FEBS Lett., 2003, 554 (3): 443–449. doi: 10.1016/S0014-5793(03)01219-5
|
| [13] |
Ghosh T K, Bian J, Gill D L. Intracellular calcium release mediated by sphingosine derivatives generated in cells. Science, 1990, 248 (4963): 1653–1656. doi: 10.1126/science.2163543
|
| [14] |
Berridge M J. Neuronal calcium signaling. Neuron, 1998, 21 (1): 13–26. doi: 10.1016/S0896-6273(00)80510-3
|
| [15] |
Grassi S, Mauri L, Prioni S, et al. Sphingosine 1-phosphate receptors and metabolic enzymes as druggable targets for brain diseases. Front. Pharmacol., 2019, 10: 807. doi: 10.3389/fphar.2019.00807
|
| [16] |
Shirakawa H, Katsumoto R, Iida S, et al. Sphingosine-1-phosphate induces Ca2+ signaling and CXCL1 release via TRPC6 channel in astrocytes. Glia, 2017, 65 (6): 1005–1016. doi: 10.1002/glia.23141
|
| [17] |
Dietrich A, Gudermann T. TRPC6: physiological function and pathophysiological relevance. Handb. Exp. Pharmacol., 2014, 222: 157–188. doi: 10.1007/978-3-642-54215-2_7
|
| [18] |
Hofmann T, Schaefer M, Schultz G, et al. Subunit composition of mammalian transient receptor potential channels in living cells. Proc. Natl. Acad. Sci. USA, 2002, 99 (11): 7461–7466. doi: 10.1073/pnas.102596199
|
| [19] |
Jeon J, Bu F, Sun G, et al. Contribution of TRPC channels in neuronal excitotoxicity associated with neurodegenerative disease and ischemic stroke. Front. Cell Dev. Biol., 2021, 8: 618663. doi: 10.3389/fcell.2020.618663
|
| [20] |
Hagenston A M, Rudnick N D, Boone C E, et al. 2-Aminoethoxydiphenyl-borate (2-APB) increases excitability in pyramidal neurons. Cell Calcium, 2009, 45 (3): 310–317. doi: 10.1016/j.ceca.2008.11.003
|
| [21] |
Sukocheva O, Wadham C, Holmes A, et al. Estrogen transactivates EGFR via the sphingosine 1-phosphate receptor Edg-3: the role of sphingosine kinase-1. J. Cell Biol., 2006, 173 (2): 301–310. doi: 10.1083/jcb.200506033
|
| [22] |
Dryer S E, Kim E Y. Permeation and rectification in canonical transient receptor potential-6 (TRPC6) channels. Front. Physiol., 2018, 9: 1055. doi: 10.3389/fphys.2018.01055
|
| [23] |
Hisatsune C, Kuroda Y, Nakamura K, et al. Regulation of TRPC6 channel activity by tyrosine phosphorylation. J. Biol. Chem., 2004, 279 (18): 18887–18894. doi: 10.1074/jbc.M311274200
|
| [24] |
Repp H, Birringer J, Koschinski A, et al. Activation of a Ca2+-dependent K+ current in mouse fibroblasts by sphingosine-1-phosphate involves the protein tyrosine kinase c-Src. Naunyn Schmiedebergs Arch. Pharmacol., 2001, 363 (3): 295–301. doi: 10.1007/s002100000362
|
| [25] |
Nodai A, Machida T, Izumi S, et al. Sphingosine 1-phosphate induces cyclooxygenase-2 via Ca2+-dependent, but MAPK-independent mechanism in rat vascular smooth muscle cells. Life Sci., 2007, 80 (19): 1768–1776. doi: 10.1016/j.lfs.2007.02.008
|
| [26] |
Berridge M J, Lipp P, Bootman M D. The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell Biol., 2000, 1 (1): 11–21. doi: 10.1038/35036035
|
| [27] |
Rapizzi E, Donati C, Cencetti F, et al. Sphingosine 1-phosphate receptors modulate intracellular Ca2+ homeostasis. Biochem. Biophys. Res. Commun., 2007, 353 (2): 268–274. doi: 10.1016/j.bbrc.2006.12.010
|
| [28] |
Chen X J, Sooch G, Demaree I S, et al. Transient Receptor Potential Canonical (TRPC) channels: Then and now. Cells, 2020, 9 (9): 1983. doi: 10.3390/cells9091983
|
| [29] |
Cheng K T, Ong H L, Liu X, et al. Contribution and regulation of TRPC channels in store-operated Ca2+ entry. Curr. Top. Membr., 2013, 71: 149–179. doi: 10.1016/B978-0-12-407870-3.00007-X
|
| [30] |
Davare M A, Fortin D A, Saneyoshi T, et al. Transient receptor potential canonical 5 channels activate Ca2+/calmodulin kinase Iγ to promote axon formation in hippocampal neurons. J. Neurosci., 2009, 29 (31): 9794–9808. doi: 10.1523/JNEUROSCI.1544-09.2009
|
| [31] |
Amaral M D, Pozzo-Miller L. TRPC3 channels are necessary for brain-derived neurotrophic factor to activate a nonselective cationic current and to induce dendritic spine formation. J. Neurosci., 2007, 27 (19): 5179–5189. doi: 10.1523/JNEUROSCI.5499-06.2007
|
| [32] |
He C K, Gao P, Cui Y T, et al. Low-glucose-sensitive TRPC6 dysfunction drives hypoglycemia-induced cognitive impairment in diabetes. Clin. Transl. Med., 2020, 10 (6): e205. doi: 10.1002/ctm2.205
|
| [33] |
Ma Y C, Huang J, Ali S, et al. Src tyrosine kinase is a novel direct effector of G proteins. Cell, 2000, 102 (5): 635–646. doi: 10.1016/S0092-8674(00)00086-6
|
| [34] |
Catarzi S, Giannoni E, Favilli F, et al. Sphingosine 1-phosphate stimulation of NADPH oxidase activity: relationship with platelet-derived growth factor receptor and c-Src kinase. Biochim. Biophys. Acta, 2007, 1770 (6): 872–883. doi: 10.1016/j.bbagen.2007.01.008
|
| [35] |
Walter D H, Rochwalsky U, Reinhold J, et al. Sphingosine-1-phosphate stimulates the functional capacity of progenitor cells by activation of the CXCR4-dependent signaling pathway via the S1P3 receptor. Arterioscler. Thromb. Vasc. Biol., 2007, 27 (2): 275–282. doi: 10.1161/01.ATV.0000254669.12675.70
|
| [36] |
Martin R, Sospedra M. Sphingosine-1 phosphate and central nervous system. Curr. Top. Microbiol. Immunol., 2014, 378: 149–170. doi: 10.1007/978-3-319-05879-5_7
|
| [37] |
Mizugishi K, Yamashita T, Olivera A, et al. Essential role for sphingosine kinases in neural and vascular development. Mol. Cell. Biol., 2005, 25 (24): 11113–11121. doi: 10.1128/MCB.25.24.11113-11121.2005
|
| [38] |
Kanno T, Nishizaki T, Proia R L, et al. Regulation of synaptic strength by sphingosine 1-phosphate in the hippocampus. Neuroscience, 2010, 171 (4): 973–980. doi: 10.1016/j.neuroscience.2010.10.021
|
| [39] |
Kajimoto T, Okada T, Yu H, et al. Involvement of sphingosine-1-phosphate in glutamate secretion in hippocampal neurons. Mol. Cell. Biol., 2007, 27 (9): 3429–3440. doi: 10.1128/MCB.01465-06
|
| [40] |
Czubowicz K, Jęśko H, Wencel P, et al. The role of ceramide and sphingosine-1-phosphate in Alzheimer’s disease and other neurodegenerative disorders. Mol. Neurobiol., 2019, 56 (8): 5436–5455. doi: 10.1007/s12035-018-1448-3
|
| [41] |
Couttas T A, Kain N, Daniels B, et al. Loss of the neuroprotective factor Sphingosine 1-phosphate early in Alzheimer’s disease pathogenesis. Acta Neuropathol. Commun., 2014, 2: 9. doi: 10.1186/2051-5960-2-9
|
| [42] |
Takasugi N, Sasaki T, Suzuki K, et al. BACE1 activity is modulated by cell-associated sphingosine-1-phosphate. J. Neurosci., 2011, 31 (18): 6850–6857. doi: 10.1523/JNEUROSCI.6467-10.2011
|
| [43] |
Malaplate-Armand C, Florent-Bechard S, Youssef I, et al. Soluble oligomers of amyloid-beta peptide induce neuronal apoptosis by activating a cPLA2-dependent sphingomyelinase-ceramide pathway. Neurobiol. Dis., 2006, 23 (1): 178–189. doi: 10.1016/j.nbd.2006.02.010
|
| [44] |
Sivasubramanian M, Kanagaraj N, Dheen S T, et al. Sphingosine kinase 2 and sphingosine-1-phosphate promotes mitochondrial function in dopaminergic neurons of mouse model of Parkinson’s disease and in MPP+-treated MN9D cells in vitro. Neuroscience, 2015, 290: 636–648. doi: 10.1016/j.neuroscience.2015.01.032
|
| [45] |
Du W L, Huang J B, Yao H L, et al. Inhibition of TRPC6 degradation suppresses ischemic brain damage in rats. J. Clin. Invest., 2010, 120 (10): 3480–3492. doi: 10.1172/JCI43165
|
| [46] |
Kim D S, Ryu H J, Kim J E, et al. The reverse roles of transient receptor potential canonical channel-3 and -6 in neuronal death following pilocarpine-induced status epilepticus. Cell. Mol. Neurobiol., 2013, 33 (1): 99–109. doi: 10.1007/s10571-012-9875-6
|
| [47] |
Tao R, Lu R, Wang J, et al. Probing the therapeutic potential of TRPC6 for Alzheimer’s disease in live neurons from patient-specific iPSCs. J. Mol. Cell Biol., 2020, 12 (10): 807–816. doi: 10.1093/jmcb/mjaa027
|
| [48] |
Lu R, Wang J, Tao R, et al. Reduced TRPC6 mRNA levels in the blood cells of patients with Alzheimer’s disease and mild cognitive impairment. Mol. Psychiatry, 2018, 23 (3): 767–776. doi: 10.1038/mp.2017.136
|
| [49] |
Wang J, Lu R, Yang J, et al. TRPC6 specifically interacts with APP to inhibit its cleavage by γ-secretase and reduce Aβ production. Nat. Commun., 2015, 6: 8876. doi: 10.1038/ncomms9876
|
| [50] |
Zhang H, Sun S, Wu L, et al. Store-operated calcium channel complex in postsynaptic spines: A new therapeutic target for Alzheimer’s disease treatment. J. Neurosci., 2016, 36 (47): 11837–11850. doi: 10.1523/JNEUROSCI.1188-16.2016
|
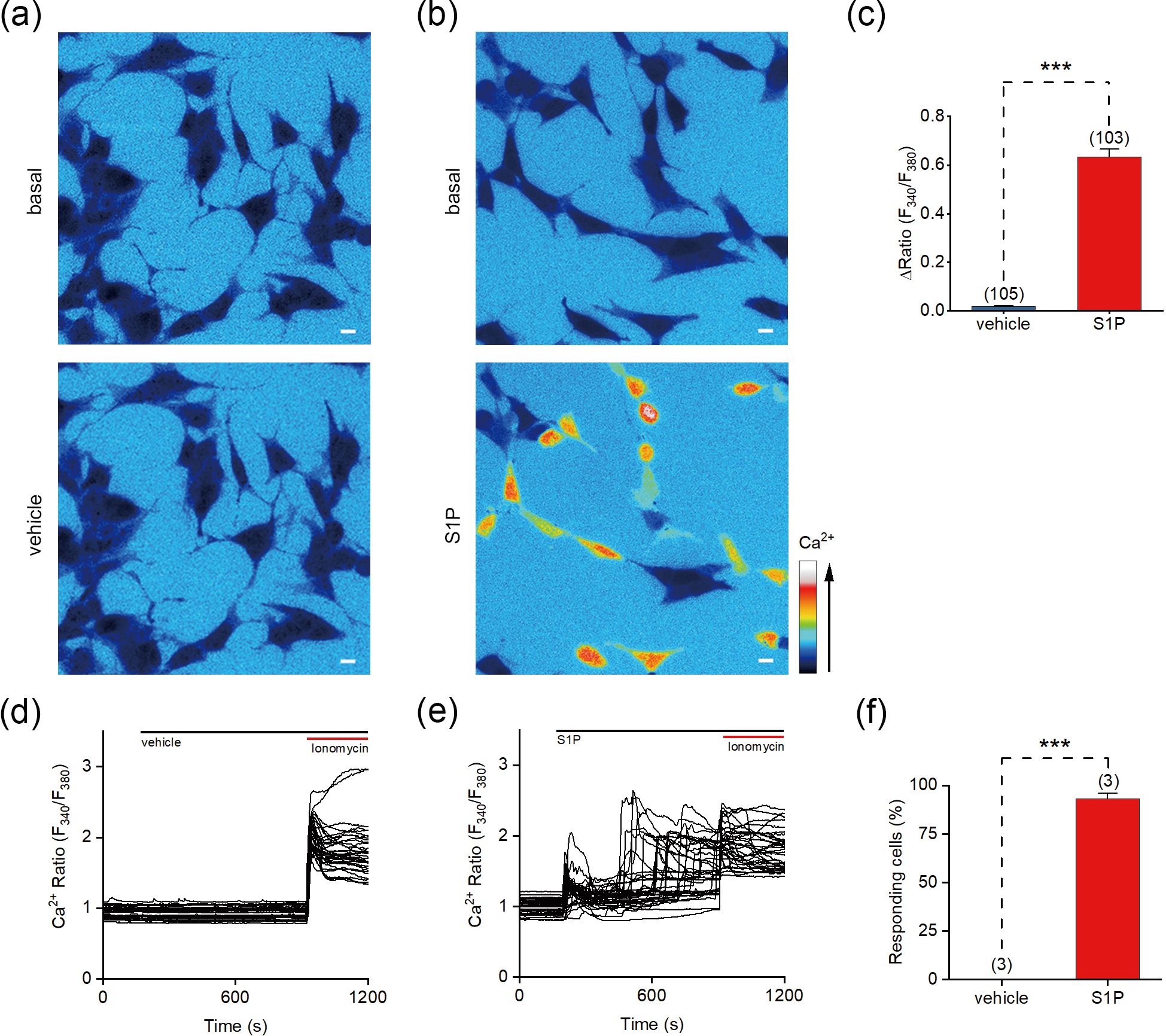
Figure 1. S1P evokes Ca2+ signals in SH-SY5Y cells. (a, b) Representative pseudocolor images of the Fura-2 AM fluorescence ratio (F340/F380) responding to vehicle (a) and 10 μmol/L S1P (b) in SH-SY5Y cells. (c) Changes in the F340/F380 ratio (ΔRatio) of cells induced by vehicle or S1P. (d, e) Representative F340/F380 ratios of SH-SY5Y cells in response to vehicle (d) or 10 µmol/L S1P (e). (f) Percentage of cells responding to the application of vehicle or S1P (ΔRatio > 0.2). Scale bar = 10 μm. The data are presented as the means ± SEMs. ***P < 0.001.
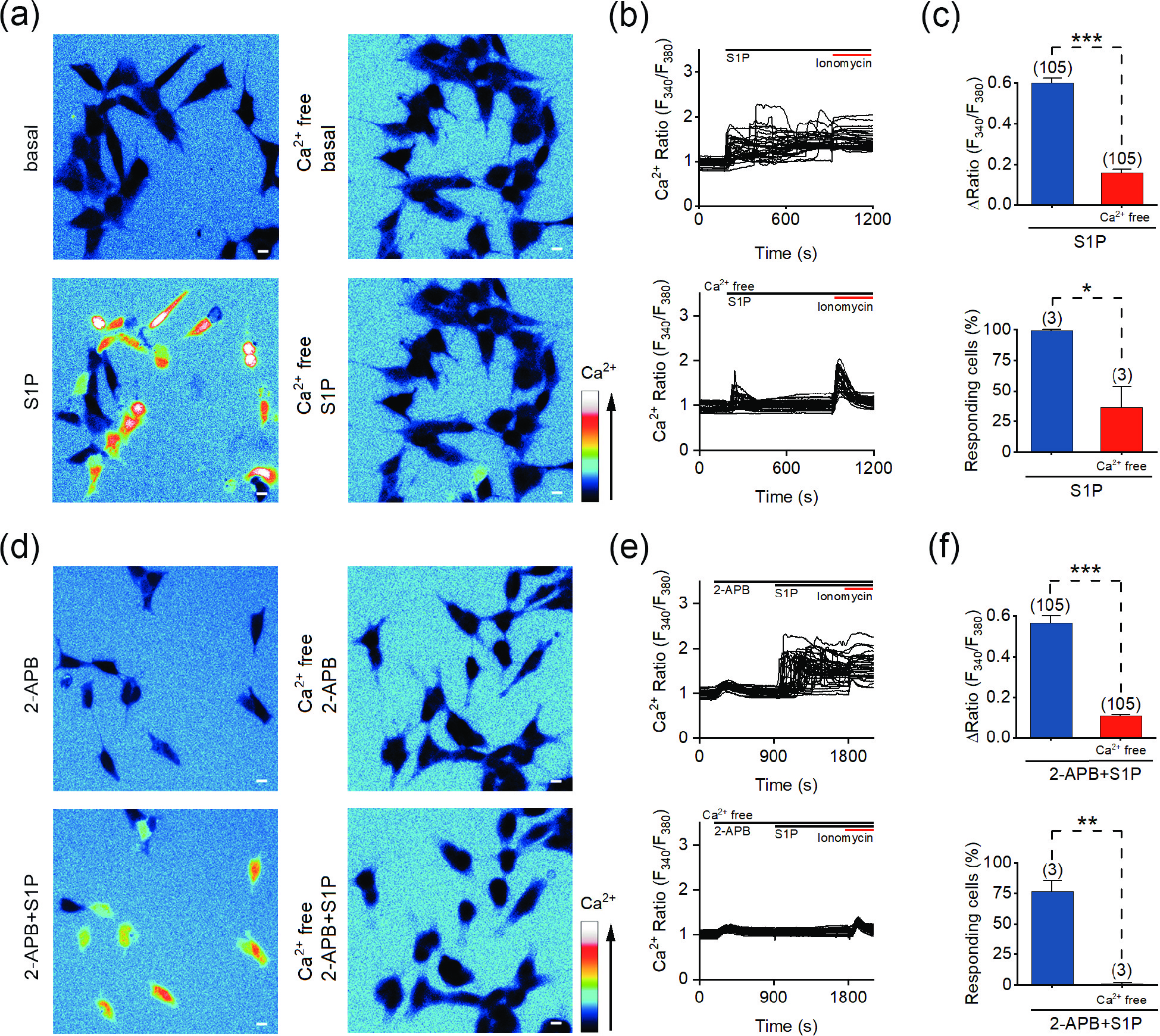
Figure 2. S1P induces the influx of extracellular Ca2+. (a) Representative pseudocolor images of F340/F380 in SH-SY5Y cells before and after the application of 10 μmol/L S1P in Ca2+-containing (left) and Ca2+-free (right) bath solution. (b) Representative traces of calcium signals responding to 10 μmol/L S1P treatment under Ca2+-containing (upper) and Ca2+-free (lower) conditions. (c) Statistics of the ΔRatio (F340/F380; upper) and percentage of responding cells (ΔRatio > 0.2) shown in (a) and (b). (d) Representative pseudocolor images of the F340/F380 ratio in SH-SY5Y cells treated with 100 μmol/L 2-APB (upper) or 100 μmol/L 2-APB + 10 μmol/L S1P (lower) in Ca2+-containing (left) and Ca2+-free (right) solution. (e) Representative F340/F380 ratios of SH-SY5Y cells in response to 100 μmol/L 2-APB + 10 μmol/L S1P in Ca2+ (upper) and Ca2+-free (lower) solution. (f) Statistics of the ΔRatio (F340/F380; upper) and percentage of responding cells (ΔRatio > 0.2) shown in (d) and (e). Scale bar = 10 μm. The data are presented as the means ± SEMs. *P < 0.05, **P < 0.01, ***P < 0.001.
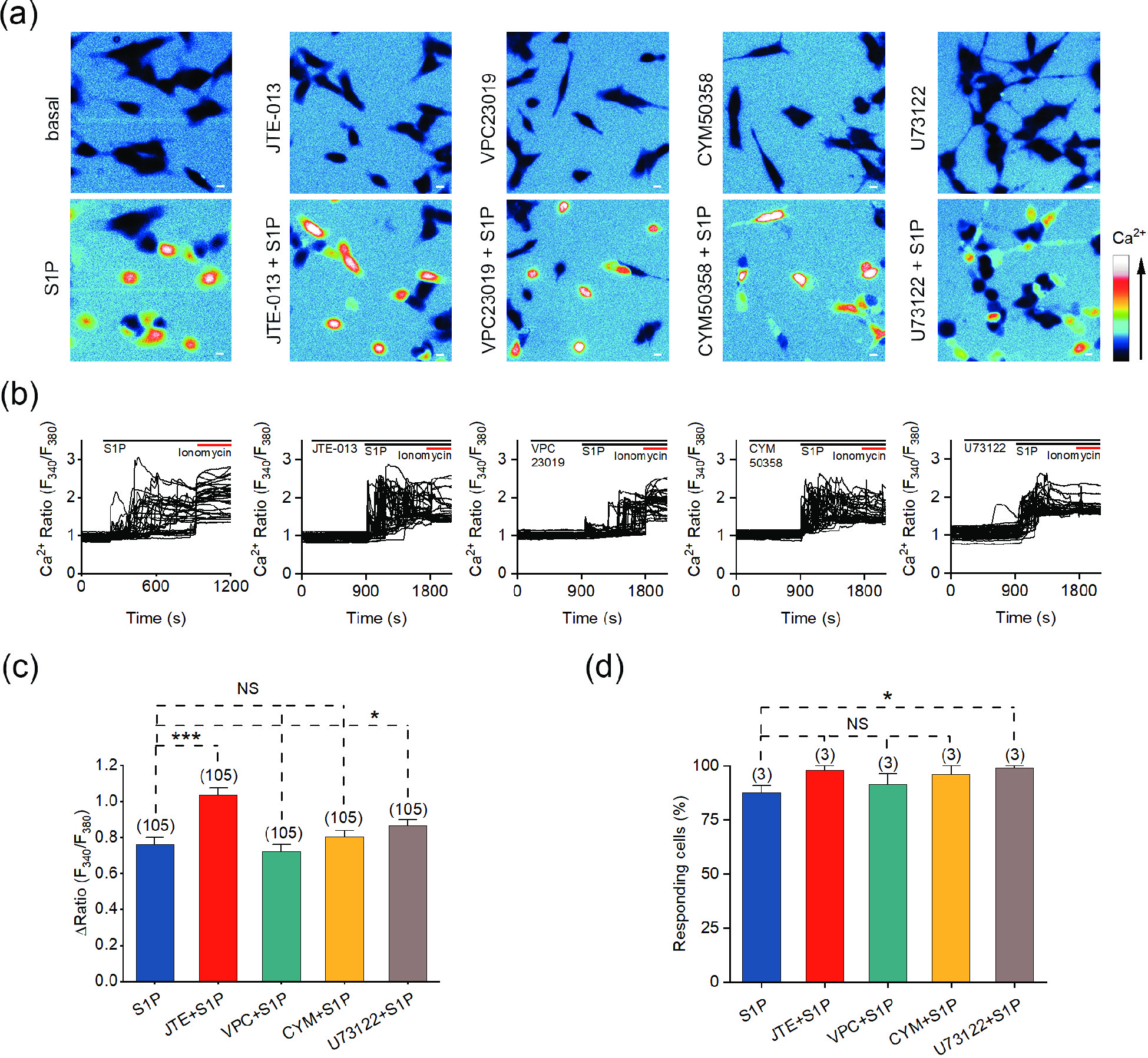
Figure 3. The S1P-induced Ca2+ response is independent of G-protein coupled S1P receptors. (a) Representative pseudocolor images of F340/F380 in SH-SY5Y cells preincubated with the indicated antagonists and treated with 10 μmol/L S1P. (b) Representative traces of calcium signals responding to 10 μmol/L S1P treatment in SH-SY5Y cells incubated with the indicated antagonists. (c) Statistics of the ΔRatio (F340/F380). (d) Statistics of the percentage of responding cells. Scale bar = 10 μm. The data are presented as the means ± SEMs. *P < 0.05, ***P < 0.001. NS indicates no significant difference.
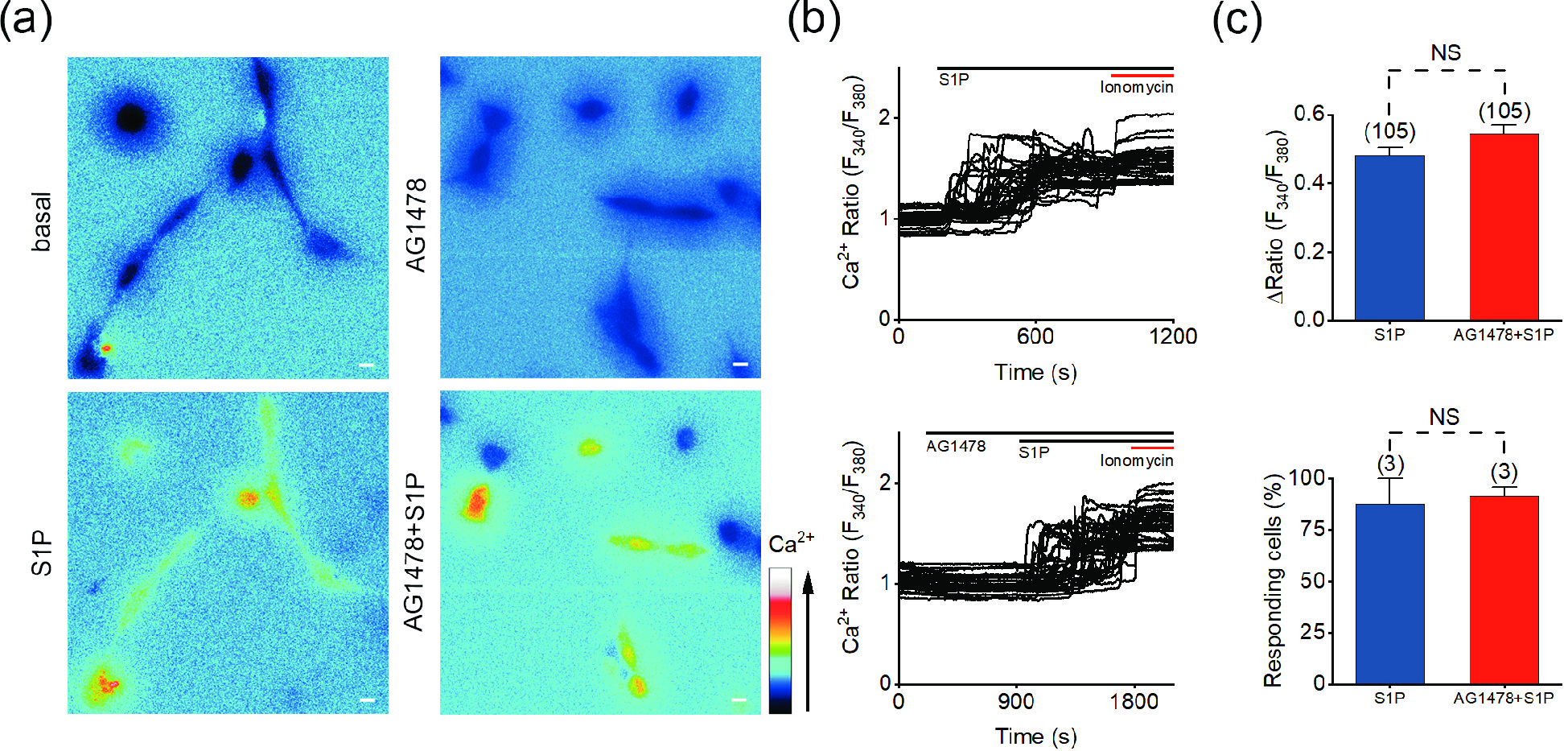
Figure 4. S1P-induced Ca2+ response is independent of the transactivation of EGFR. (a) Representative pseudocolor images of F340/F380 in SH-SY5Y cells preincubated with 1 μmol/L AG1478 and treated with 10 μmol/L S1P. (b) Representative traces of calcium signals responding to 10 μmol/L S1P treatment in SH-SY5Y cells incubated with 1 μmol/L AG1478. (c) Statistics of the ΔRatio (F340/F380) and statistics of the percentage of responding cells. Scale bar = 10 μm. The data are presented as the means ± SEMs. NS indicates no significant difference.
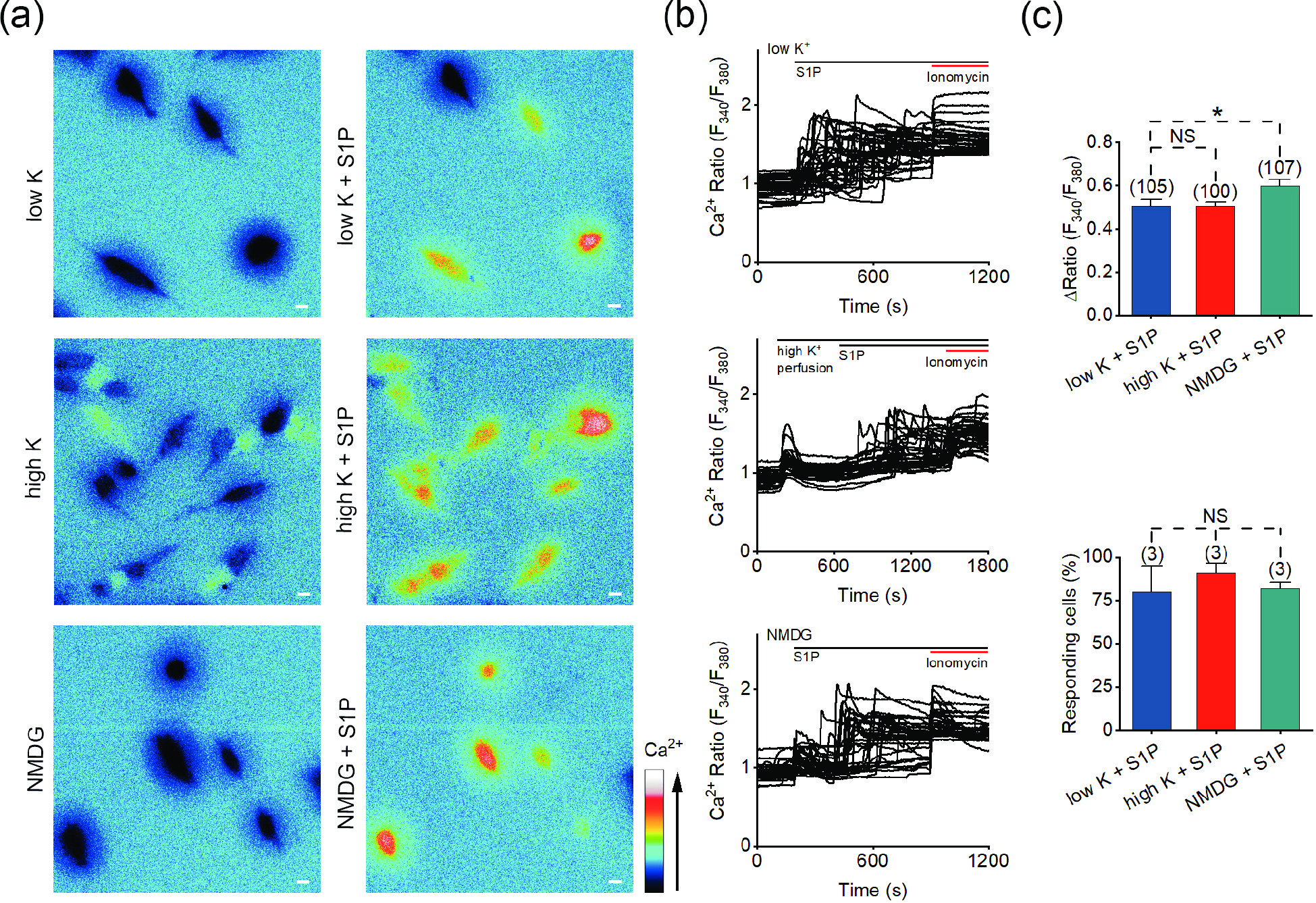
Figure 5. S1P induces calcium signals in high-K+ solution and Na+-free solution. (a) Representative pseudocolor images of F340/F380 showing the S1P-induced calcium signals in SH-SY5Y cells with low-K+ (5.4 mmol/L; upper), high-K+ (100 mmol/L; middle) or Na+-free (lower) solution treatment. (b) Representative traces of calcium signals responding to low-K+ (upper), high-K+ (middle) or Na+-free (lower) treatment in SH-SY5Y cells. (c) Statistics of the ΔRatio (F340/F380; upper) and percentage of responding cells (ΔRatio > 0.2; lower) shown in (a) and (b). Scale bar = 10 μm. The data are presented as the means ± SEMs. *P < 0.05. NS indicates no significant difference.

Figure 6. S1P-induced Ca2+ mobilization is mediated by the TRPC6 channel in both SH-SY5Y cells and hippocampal neurons. (a) Representative pseudocolor images of F340/F380 showing the S1P-induced calcium signals in SH-SY5Y cells preincubated with vehicle (left) or 1 μmol/L SAR7334 (right). (b) Representative traces of calcium signals responding to 10 μmol/L S1P treatment in SH-SY5Y cells preincubated with vehicle (upper) or 1 μmol/L SAR7334 (lower). (c) Statistics of the ΔRatio (F340/F380; upper) and percentage of responding cells (ΔRatio > 0.2) shown in (a) and (b). (d) Representative pseudocolor images of F340/F380 showing the S1P-induced calcium signals in hippocampal neurons preincubated with vehicle (left) or 0.1 μmol/L SAR7334 (right). (e) Representative traces of calcium signals responding to 10 μmol/L S1P treatment in hippocampal neurons preincubated with vehicle (upper) or 0.1 μmol/L SAR7334 (lower). (f) Statistics of the ΔRatio (F340/F380; upper) and percentage of responding cells (ΔRatio > 0.2) shown in (c) and (d). Scale bar = 10 μm. The data are presented as the means ± SEMs. **P < 0.01, ***P < 0.001.
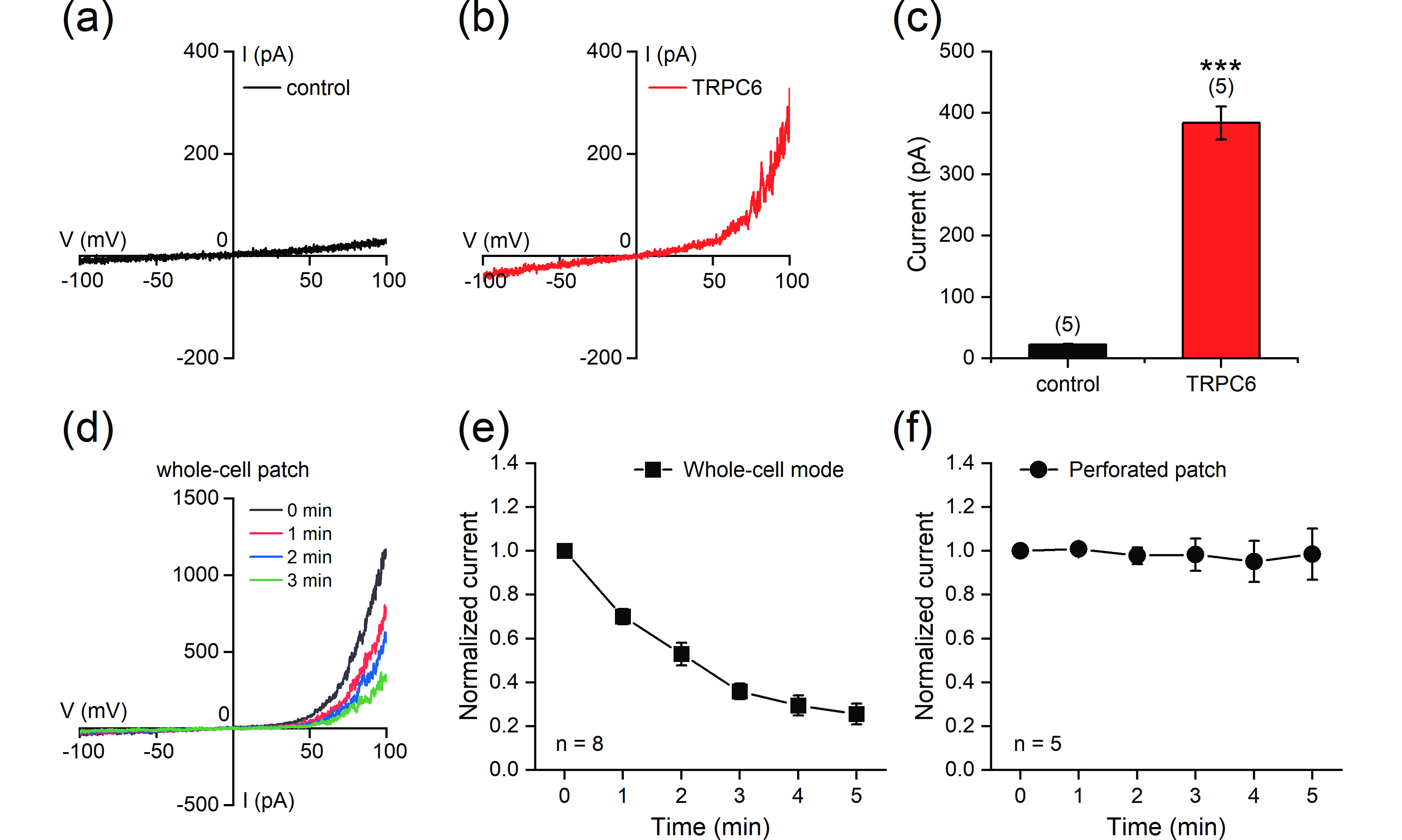
Figure 7. Reconstitution of the TRPC6 current in HEK293T cells. (a, b) Representative whole-cell currents recorded in HEK293T cells transfected with empty vector (a) or TRPC6 (b). The currents were elicited with ramp protocols (−100 mV to +100 mV in 1 s). (c) Current amplitudes measured at +100 mV. (d) Representative currents recorded using whole-cell patch-clamp in TRPC6-transfected HEK293T cells at different time points after the start of recording. (e, f) Stability of the currents under whole-cell patch-clamp mode (e) or perforated patch-clamp mode (f). The current amplitudes (measured at +100 mV) at different time points were normalized to the values at the start of recordings. The data are presented as the means ± SEMs. ***P < 0.001.
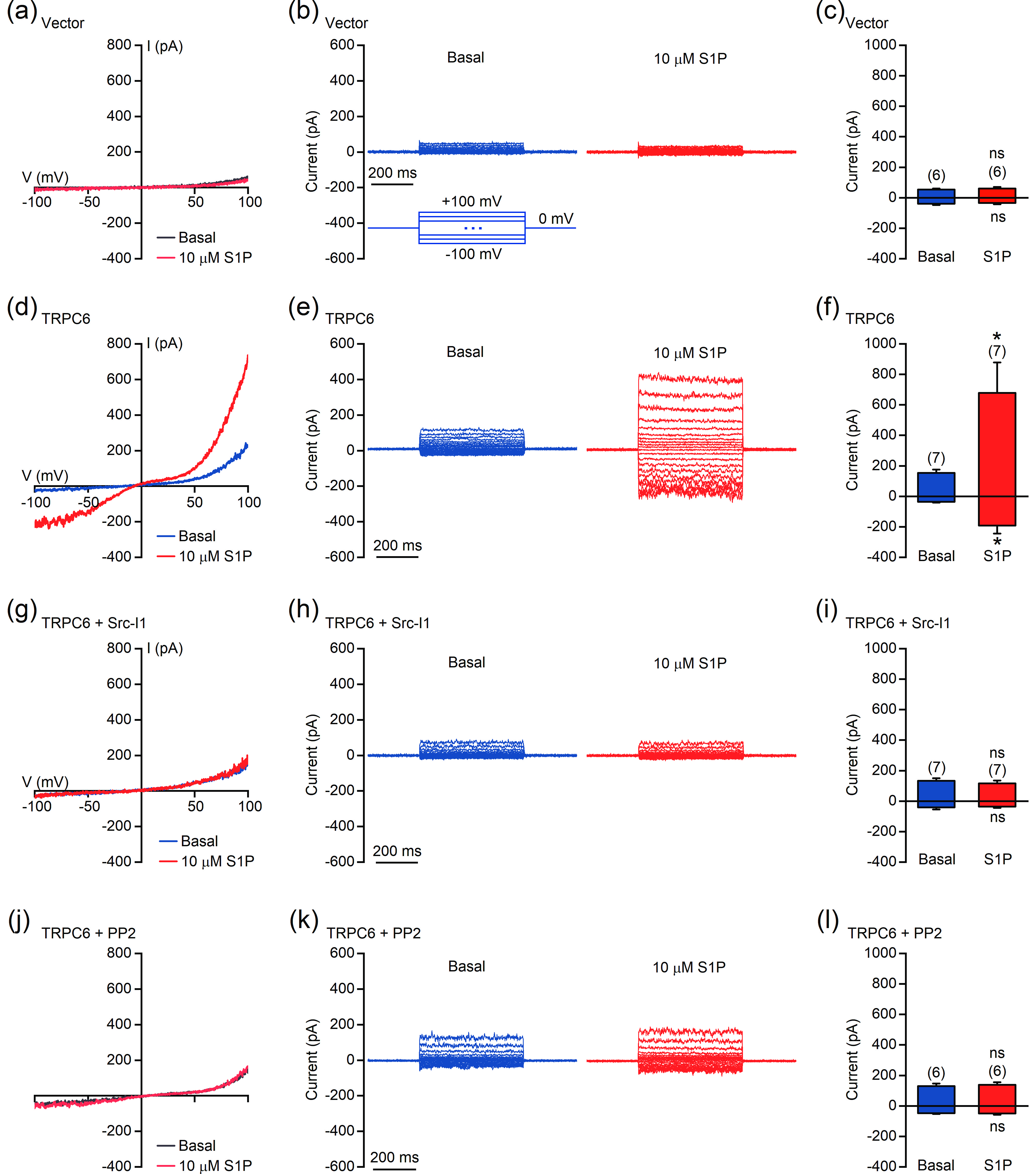
Figure 8. S1P activates TRPC6 in a Src-dependent manner. (a–l) Perforated patch-clamp recordings were performed in HEK293T cells transfected with empty vector (a–c) or TRPC6 (d–i). Cells used in (g–l) were treated with 2 μmol/L Src-I1 (g–i) or 1 μmol/L PP2 (j–l) for 10 min before recording. (a, d, g, and j) Representative currents recorded with ramp protocols (−100 to +100 mV in 1 s). (b, e, h, and k) Representative currents recorded with step protocols (250 ms step pulses from −100 to +100 mV, 10 mV step; Vh = 0 mV). (c, f, i, and l) Amplitudes of the inward and outward currents shown in (a), (d), (g), and (j), respectively. The data are presented as the means ± SEMs. *P < 0.05. NS indicates no significant difference.
| [1] |
Dixit D, Okuniewska M, Schwab S R. Secrets and lyase: Control of sphingosine 1-phosphate distribution. Immunol. Rev., 2019, 289 (1): 173–185. doi: 10.1111/imr.12760
|
| [2] |
Książek M, Chacińska M, Chabowski A, et al. Sources, metabolism, and regulation of circulating sphingosine-1-phosphate. J. Lipid Res., 2015, 56 (7): 1271–1281. doi: 10.1194/jlr.R059543
|
| [3] |
Spiegel S, Milstien S. Sphingosine-1-phosphate: an enigmatic signalling lipid. Nat. Rev. Mol. Cell Biol., 2003, 4 (5): 397–407. doi: 10.1038/nrm1103
|
| [4] |
Nagahashi M, Takabe K, Terracina K P, et al. Sphingosine-1-phosphate transporters as targets for cancer therapy. Biomed Res. Int., 2014, 2014: 651727. doi: 10.1155/2014/651727
|
| [5] |
Chun J, Hla T, Lynch K R, et al. International Union of Basic and Clinical Pharmacology. LXXVIII. Lysophospholipid receptor nomenclature. Pharmacol. Rev., 2010, 62 (4): 579–587. doi: 10.1124/pr.110.003111
|
| [6] |
Hait N C, Oskeritzian C A, Paugh S W, et al. Sphingosine kinases, sphingosine 1-phosphate, apoptosis and diseases. Biochim. Biophys. Acta, 2006, 1758 (12): 2016–2026. doi: 10.1016/j.bbamem.2006.08.007
|
| [7] |
Ng M L, Yarla N S, Menschikowski M, et al. Regulatory role of sphingosine kinase and sphingosine-1-phosphate receptor signaling in progenitor/stem cells. World J. Stem Cells, 2018, 10 (9): 119–133. doi: 10.4252/wjsc.v10.i9.119
|
| [8] |
Sukocheva O A. Expansion of sphingosine kinase and sphingosine-1-phosphate receptor function in normal and cancer cells: From membrane restructuring to mediation of estrogen signaling and stem cell programming. Int. J. Mol. Sci., 2018, 19 (2): 420. doi: 10.3390/ijms19020420
|
| [9] |
Birchwood C J, Saba J D, Dickson R C, et al. Calcium influx and signaling in yeast stimulated by intracellular sphingosine 1-phosphate accumulation. J. Biol. Chem., 2001, 276 (15): 11712–11718. doi: 10.1074/jbc.M010221200
|
| [10] |
Pulli I, Asghar M Y, Kemppainen K, et al. Sphingolipid-mediated calcium signaling and its pathological effects. Biochim. Biophys. Acta Mol. Cell Res., 2018, 1865 (11 Pt B): 1668–1677. doi: 10.1016/j.bbamcr.2018.04.012
|
| [11] |
Putney J W, Tomita T. Phospholipase C signaling and calcium influx. Adv. Biol. Regul., 2012, 52 (1): 152–164. doi: 10.1016/j.advenzreg.2011.09.005
|
| [12] |
Meyer zu Heringdorf D, Liliom K, Schaefer M, et al. Photolysis of intracellular caged sphingosine-1-phosphate causes Ca2+ mobilization independently of G-protein-coupled receptors. FEBS Lett., 2003, 554 (3): 443–449. doi: 10.1016/S0014-5793(03)01219-5
|
| [13] |
Ghosh T K, Bian J, Gill D L. Intracellular calcium release mediated by sphingosine derivatives generated in cells. Science, 1990, 248 (4963): 1653–1656. doi: 10.1126/science.2163543
|
| [14] |
Berridge M J. Neuronal calcium signaling. Neuron, 1998, 21 (1): 13–26. doi: 10.1016/S0896-6273(00)80510-3
|
| [15] |
Grassi S, Mauri L, Prioni S, et al. Sphingosine 1-phosphate receptors and metabolic enzymes as druggable targets for brain diseases. Front. Pharmacol., 2019, 10: 807. doi: 10.3389/fphar.2019.00807
|
| [16] |
Shirakawa H, Katsumoto R, Iida S, et al. Sphingosine-1-phosphate induces Ca2+ signaling and CXCL1 release via TRPC6 channel in astrocytes. Glia, 2017, 65 (6): 1005–1016. doi: 10.1002/glia.23141
|
| [17] |
Dietrich A, Gudermann T. TRPC6: physiological function and pathophysiological relevance. Handb. Exp. Pharmacol., 2014, 222: 157–188. doi: 10.1007/978-3-642-54215-2_7
|
| [18] |
Hofmann T, Schaefer M, Schultz G, et al. Subunit composition of mammalian transient receptor potential channels in living cells. Proc. Natl. Acad. Sci. USA, 2002, 99 (11): 7461–7466. doi: 10.1073/pnas.102596199
|
| [19] |
Jeon J, Bu F, Sun G, et al. Contribution of TRPC channels in neuronal excitotoxicity associated with neurodegenerative disease and ischemic stroke. Front. Cell Dev. Biol., 2021, 8: 618663. doi: 10.3389/fcell.2020.618663
|
| [20] |
Hagenston A M, Rudnick N D, Boone C E, et al. 2-Aminoethoxydiphenyl-borate (2-APB) increases excitability in pyramidal neurons. Cell Calcium, 2009, 45 (3): 310–317. doi: 10.1016/j.ceca.2008.11.003
|
| [21] |
Sukocheva O, Wadham C, Holmes A, et al. Estrogen transactivates EGFR via the sphingosine 1-phosphate receptor Edg-3: the role of sphingosine kinase-1. J. Cell Biol., 2006, 173 (2): 301–310. doi: 10.1083/jcb.200506033
|
| [22] |
Dryer S E, Kim E Y. Permeation and rectification in canonical transient receptor potential-6 (TRPC6) channels. Front. Physiol., 2018, 9: 1055. doi: 10.3389/fphys.2018.01055
|
| [23] |
Hisatsune C, Kuroda Y, Nakamura K, et al. Regulation of TRPC6 channel activity by tyrosine phosphorylation. J. Biol. Chem., 2004, 279 (18): 18887–18894. doi: 10.1074/jbc.M311274200
|
| [24] |
Repp H, Birringer J, Koschinski A, et al. Activation of a Ca2+-dependent K+ current in mouse fibroblasts by sphingosine-1-phosphate involves the protein tyrosine kinase c-Src. Naunyn Schmiedebergs Arch. Pharmacol., 2001, 363 (3): 295–301. doi: 10.1007/s002100000362
|
| [25] |
Nodai A, Machida T, Izumi S, et al. Sphingosine 1-phosphate induces cyclooxygenase-2 via Ca2+-dependent, but MAPK-independent mechanism in rat vascular smooth muscle cells. Life Sci., 2007, 80 (19): 1768–1776. doi: 10.1016/j.lfs.2007.02.008
|
| [26] |
Berridge M J, Lipp P, Bootman M D. The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell Biol., 2000, 1 (1): 11–21. doi: 10.1038/35036035
|
| [27] |
Rapizzi E, Donati C, Cencetti F, et al. Sphingosine 1-phosphate receptors modulate intracellular Ca2+ homeostasis. Biochem. Biophys. Res. Commun., 2007, 353 (2): 268–274. doi: 10.1016/j.bbrc.2006.12.010
|
| [28] |
Chen X J, Sooch G, Demaree I S, et al. Transient Receptor Potential Canonical (TRPC) channels: Then and now. Cells, 2020, 9 (9): 1983. doi: 10.3390/cells9091983
|
| [29] |
Cheng K T, Ong H L, Liu X, et al. Contribution and regulation of TRPC channels in store-operated Ca2+ entry. Curr. Top. Membr., 2013, 71: 149–179. doi: 10.1016/B978-0-12-407870-3.00007-X
|
| [30] |
Davare M A, Fortin D A, Saneyoshi T, et al. Transient receptor potential canonical 5 channels activate Ca2+/calmodulin kinase Iγ to promote axon formation in hippocampal neurons. J. Neurosci., 2009, 29 (31): 9794–9808. doi: 10.1523/JNEUROSCI.1544-09.2009
|
| [31] |
Amaral M D, Pozzo-Miller L. TRPC3 channels are necessary for brain-derived neurotrophic factor to activate a nonselective cationic current and to induce dendritic spine formation. J. Neurosci., 2007, 27 (19): 5179–5189. doi: 10.1523/JNEUROSCI.5499-06.2007
|
| [32] |
He C K, Gao P, Cui Y T, et al. Low-glucose-sensitive TRPC6 dysfunction drives hypoglycemia-induced cognitive impairment in diabetes. Clin. Transl. Med., 2020, 10 (6): e205. doi: 10.1002/ctm2.205
|
| [33] |
Ma Y C, Huang J, Ali S, et al. Src tyrosine kinase is a novel direct effector of G proteins. Cell, 2000, 102 (5): 635–646. doi: 10.1016/S0092-8674(00)00086-6
|
| [34] |
Catarzi S, Giannoni E, Favilli F, et al. Sphingosine 1-phosphate stimulation of NADPH oxidase activity: relationship with platelet-derived growth factor receptor and c-Src kinase. Biochim. Biophys. Acta, 2007, 1770 (6): 872–883. doi: 10.1016/j.bbagen.2007.01.008
|
| [35] |
Walter D H, Rochwalsky U, Reinhold J, et al. Sphingosine-1-phosphate stimulates the functional capacity of progenitor cells by activation of the CXCR4-dependent signaling pathway via the S1P3 receptor. Arterioscler. Thromb. Vasc. Biol., 2007, 27 (2): 275–282. doi: 10.1161/01.ATV.0000254669.12675.70
|
| [36] |
Martin R, Sospedra M. Sphingosine-1 phosphate and central nervous system. Curr. Top. Microbiol. Immunol., 2014, 378: 149–170. doi: 10.1007/978-3-319-05879-5_7
|
| [37] |
Mizugishi K, Yamashita T, Olivera A, et al. Essential role for sphingosine kinases in neural and vascular development. Mol. Cell. Biol., 2005, 25 (24): 11113–11121. doi: 10.1128/MCB.25.24.11113-11121.2005
|
| [38] |
Kanno T, Nishizaki T, Proia R L, et al. Regulation of synaptic strength by sphingosine 1-phosphate in the hippocampus. Neuroscience, 2010, 171 (4): 973–980. doi: 10.1016/j.neuroscience.2010.10.021
|
| [39] |
Kajimoto T, Okada T, Yu H, et al. Involvement of sphingosine-1-phosphate in glutamate secretion in hippocampal neurons. Mol. Cell. Biol., 2007, 27 (9): 3429–3440. doi: 10.1128/MCB.01465-06
|
| [40] |
Czubowicz K, Jęśko H, Wencel P, et al. The role of ceramide and sphingosine-1-phosphate in Alzheimer’s disease and other neurodegenerative disorders. Mol. Neurobiol., 2019, 56 (8): 5436–5455. doi: 10.1007/s12035-018-1448-3
|
| [41] |
Couttas T A, Kain N, Daniels B, et al. Loss of the neuroprotective factor Sphingosine 1-phosphate early in Alzheimer’s disease pathogenesis. Acta Neuropathol. Commun., 2014, 2: 9. doi: 10.1186/2051-5960-2-9
|
| [42] |
Takasugi N, Sasaki T, Suzuki K, et al. BACE1 activity is modulated by cell-associated sphingosine-1-phosphate. J. Neurosci., 2011, 31 (18): 6850–6857. doi: 10.1523/JNEUROSCI.6467-10.2011
|
| [43] |
Malaplate-Armand C, Florent-Bechard S, Youssef I, et al. Soluble oligomers of amyloid-beta peptide induce neuronal apoptosis by activating a cPLA2-dependent sphingomyelinase-ceramide pathway. Neurobiol. Dis., 2006, 23 (1): 178–189. doi: 10.1016/j.nbd.2006.02.010
|
| [44] |
Sivasubramanian M, Kanagaraj N, Dheen S T, et al. Sphingosine kinase 2 and sphingosine-1-phosphate promotes mitochondrial function in dopaminergic neurons of mouse model of Parkinson’s disease and in MPP+-treated MN9D cells in vitro. Neuroscience, 2015, 290: 636–648. doi: 10.1016/j.neuroscience.2015.01.032
|
| [45] |
Du W L, Huang J B, Yao H L, et al. Inhibition of TRPC6 degradation suppresses ischemic brain damage in rats. J. Clin. Invest., 2010, 120 (10): 3480–3492. doi: 10.1172/JCI43165
|
| [46] |
Kim D S, Ryu H J, Kim J E, et al. The reverse roles of transient receptor potential canonical channel-3 and -6 in neuronal death following pilocarpine-induced status epilepticus. Cell. Mol. Neurobiol., 2013, 33 (1): 99–109. doi: 10.1007/s10571-012-9875-6
|
| [47] |
Tao R, Lu R, Wang J, et al. Probing the therapeutic potential of TRPC6 for Alzheimer’s disease in live neurons from patient-specific iPSCs. J. Mol. Cell Biol., 2020, 12 (10): 807–816. doi: 10.1093/jmcb/mjaa027
|
| [48] |
Lu R, Wang J, Tao R, et al. Reduced TRPC6 mRNA levels in the blood cells of patients with Alzheimer’s disease and mild cognitive impairment. Mol. Psychiatry, 2018, 23 (3): 767–776. doi: 10.1038/mp.2017.136
|
| [49] |
Wang J, Lu R, Yang J, et al. TRPC6 specifically interacts with APP to inhibit its cleavage by γ-secretase and reduce Aβ production. Nat. Commun., 2015, 6: 8876. doi: 10.1038/ncomms9876
|
| [50] |
Zhang H, Sun S, Wu L, et al. Store-operated calcium channel complex in postsynaptic spines: A new therapeutic target for Alzheimer’s disease treatment. J. Neurosci., 2016, 36 (47): 11837–11850. doi: 10.1523/JNEUROSCI.1188-16.2016
|

ISSN 0253-2778
CN 34-1054/N
Copyright © Editorial Office of JUSTC, All Rights Reserved. 皖ICP备05002528号
Supported by:
Beijing Renhe Information Technology Co. Ltd





 DownLoad:
DownLoad: